Impact Factor ISSN: 1449-1907
- Issue 9; 2026
- Issue 8; 2026
- Issue 7; 2026
- Issue 6; 2026
- Issue 5; 2026
- Volume 23; 2026
- Past Issues
- Editorial Board
- Cover Images
- Index & Coverage
- Cover Suggestion
- Special Issues
Introduction
Materials and methods
Results
Discussion
Supplementary Material
Acknowledgements
References

 Global reach, higher impact
Global reach, higher impactInt J Med Sci 2021; 18(6):1339-1347. doi:10.7150/ijms.49997 This issue Cite
Research Paper
Research on the circadian clock gene HNF4a in different malignant tumors
Meng-jun Qiu1, Li Zhang1, Xie-fan Fang2, Qiu-ting Li1, Li-sheng Zhu3, Bin Zhang3, Sheng-li Yang3 ![]() , Zhi-fan Xiong1
, Zhi-fan Xiong1 ![]()
1. Division of Gastroenterology, Liyuan Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430077, China.
2. Charles River Laboratories, Inc., 6995 Longley Lane, Reno NV 89511.
3. Cancer Center, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China.
Received 2020-6-27; Accepted 2021-1-4; Published 2021-1-21
Abstract

Background: The circadian rhythm is produced by multiple feedback loops formed by the core clock genes after transcription and translation, thus regulating various metabolic and physiological functions of the human body. We have shown previously that the abnormal expression of 14 clock genes is related closely to the occurrence and development of different malignant tumors, and these genes may play an anti-cancer or pro-cancer role in different tumors. HNF4a has many typical properties of clock proteins involved in the clock gene negative feedback loop regulation process. We need to explore the function of HNF4a as a circadian clock gene in malignant tumors further. Methods: We used The Cancer Genome Atlas (TCGA) database to download the clinicopathological information of twenty malignant tumors and the corresponding RNA-seq data. The HNF4a RNA-seq data standardized by R language and clinical information were integrated to reveal the relationship between HNF4a and prognosis of patients. Results: Analysis of TCGA data showed that the prognosis of HNF4a was significantly different in BLCA, KIRC, LUSC, and READ. High HNF4a expression is correlated with good prognosis in BLCA, KIRC, and READ but poor prognosis in LUSC. However, HNF4a was associated with the stages, T stages, and lymph node status only in BLCA. Conclusions: HNF4a plays different roles in different malignancies, and the abnormal expression of HNF4a has a great correlation with the biological characteristics of BLCA. The low expression of HNF4a could be a reference index for the metastasis, recurrence, and prognosis of BLCA.
Keywords: circadian clock, HNF4a, malignant tumors, prognosis.
Introduction
HNF4a is a member of the nuclear receptor superfamily HNF4, and is also a hepatic cell-specific transcription factor. In addition to its expression in the liver, HNF4a is commonly expressed in organs containing epithelial cells, whose transcriptional translation involves a regulated expression of many activity-related target genes, such as lipid transport, glucose, cholesterol, amino acid metabolism, and the regulation of the blood system [1-4]. Changes in physiological activity, such as changes in insulin levels after a meal and changes in the products of dietary metabolisms, such as bile acids, can affect the expression level of HNF4a [5-7]. The importance of maintaining physiological homeostasis is reflected in the function of HNF4a in drug metabolism and its potential as a drug target [8].
The poor differentiation and immortalization of cells are the bases of tumorigenesis. It has been shown that HNF4a can not only reduce or inhibit cell proliferation but can also accelerate the differentiation speed of poorly differentiated liver cancer by re-expressing HNF4a in liver cancer and maintaining a highly differentiated phenotype, which reduces its invasiveness [9-11]. Cell growth includes proliferation and apoptosis, and the comprehensive effects of the two are ultimately manifested as the strength of cell growth ability. A cell cycle closely related to apoptosis is also an important factor in tumor development, and the overexpression of HNF4a has been found to inhibit G1 cell cycle progression in reported work [12].
Circadian rhythm is a physiological, biochemical, and behavioral activity that organisms used to adapt to periodic changes in the environment [13]. The basic working principle of this activity relies mainly on the regulation of the transcription of the core clock element to achieve the consistency of the rhythm of each organ of the body with the circadian rhythm of the external environment [14]. At the molecular level, maintaining the normal function of the circadian rhythm involves the negative feedback loop formed by a series of clock genes, namely the transcription/translation negative feedback, which regulates circadian rhythms at the cellular level, governing sleep, eating, temperature, hormonal secretion, and a number of metabolic pathways [14, 15]. The disruption of the normal circadian rhythm can lead to clinically relevant diseases, including cardiovascular diseases, neurodegenerative diseases, diabetes, and various cancers [16-20].
HNF4a is a clock gene that is mainly studied in the field of liver cancer. It can play an anti-cancer role by inhibiting the initiation, growth of hepatocellular carcinoma, or progressions of some genes, such as cyclin D1 and CCND1 [9, 21-23], and act as a transcriptional activator, interacting with repressor proteins to allow the inhibition of some target genes [24-26]. The gene can also affect apoptosis and cell cycle by upregulating Exo70 and inhibiting the epithelial-mesenchymal transformation gene or inhibit the expression of the circadian protein BMAL1 directly [27, 28]. The exact association between HNF4a and cancers remains unclear. In this work, the TCGA database was used to analyze the expression of HNF4a in twenty malignant tumors (adrenocortical carcinoma (ACC), bladder urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA), cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC), cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), lymphoid neoplasm diffuse large B-cell Lymphoma (DLBC), esophageal carcinoma (ESCA), head and neck squamous cell carcinoma (HNSC), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), ovarian serous cystadenocarcinoma (OV), pancreatic adenocarcinoma (PAAD), rectum adenocarcinoma (READ), sarcoma (SARC), stomach adenocarcinoma (STAD), uterine corpus endometrial carcinoma (UCEC)) to evaluate the patient prognoses and clinicopathological features.
Materials and methods
The RNA-Seq of HNF4a in twenty different malignant tumors and the corresponding clinical data were downloaded from the TCGA website (https://cancergenome.nih.gov/). The RNA-Seq data from 79 ACC tissues, 408 BLCA tissues, 1093 BRCA tissues, 304 CESC tissues, 36 CHOL tissues, 457 COAD tissues, 48 DLBC tissues, 184 ESCA tissues, 520 HNSC tissues, 533 KIRC tissues, 290 KIRP tissues, 371 LIHC tissues, 515 LUAD tissues, 501 LUSC tissues, 304 OV tissues, 178 PAAD tissues, 105 READ tissues, 259 SARC tissues, 415 STAD tissues, and 545 UCEC tissues. In this study, we integrated the RNA-Seq data standardized by the software edgeR from Bioconductor (http://www.bioconductor.org/) and clinical data according to the serial number of each patient and specimen provided by TCGA. Since the above two types of data are not exactly matched, we performed the culling of the mismatched data. Finally, the analysis of the prognoses and clinicopathological data of HNF4a in twenty different malignant tumors was performed on the data from 79 ACC tissues, 405 BLCA tissues, 1092 BRCA tissues, 304 CESC tissues, 36 CHOL tissues, 298 COAD tissues, 48 DLBC tissues, 184 ESCA tissues, 519 HNSC tissues, 533 KIRC tissues, 289 KIRP tissues, 370 LIHC tissues, 507 LUAD tissues, 495 LUSC tissues, 303 OV tissues, 178 PAAD tissues, 97 READ tissues, 259 SARC tissues, 408 STAD tissues, and 181 UCEC tissues. We assessed the relationship between the differential expression of HNF4a according to the median of the data and survival time and finally made the survival curve. We also examined the correlation between HNF4a expression and associated clinicopathological information with a variable degree of deletion in those cancers which has significantly different prognosis. Finally, we used the Genotype-Tissue Expressions (GTEx) database (http://commonfund.nih.gov/GTEx/) to analyze the expression differences of HNF4a in various normal tissues. Statistical analysis was performed with the SPSS® version 18.0. The variance analysis of the effects of HNF4a on prognosis was performed using Kaplan-Meier survival curves and the log-rank test. The indexes between the HNF4a expression and clinicopathological information, as well as the correlation between HNF4a and other clock genes, were compared using Pearson's Chi-square test.
Results
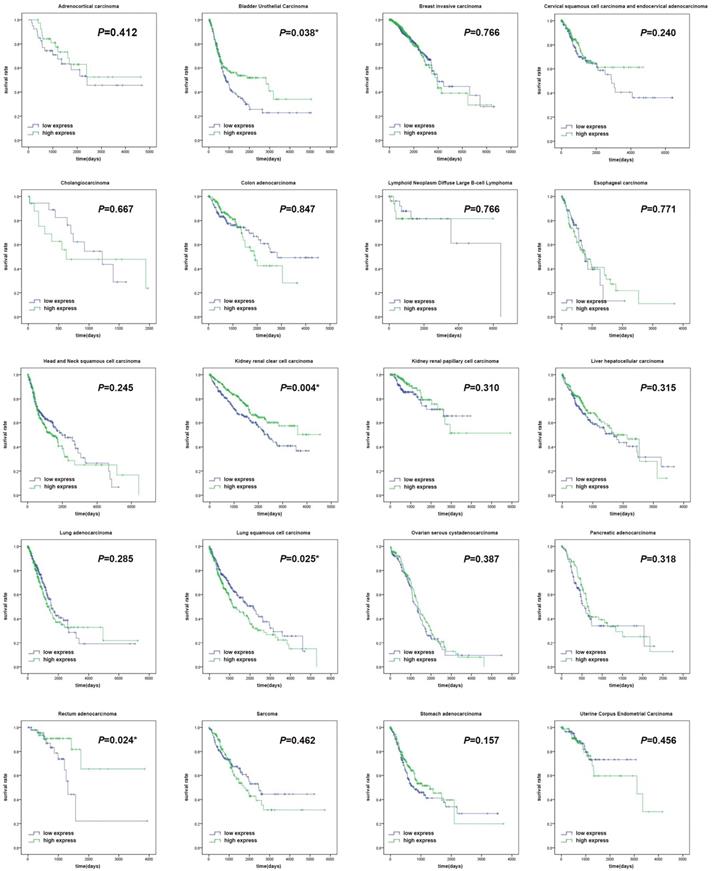
The expression levels of HNF4a in the twenty different malignant tumors were related to different prognoses. It was shown in our study that patients with high HNF4a expression showed a significantly longer overall survival (OS, p < 0.05) than patients with low HNF4a expression in BLCA, KIRC, and READ, while patients with high expression of HNF4a exhibited a significantly shorter OS (p < 0.05) than patients with low expression of HNF4a in LUSC (Figure 1). There were no significant differences in other malignant tumors (p > 0.05). We extracted the mRNA levels of HNF4a from some normal tissues in GTEx and plotted a box diagram of the HNF4a expression. As shown in Supplementary Figure 1, HNF4a mRNA levels increased relatively in kidney, bladder, colorectum, liver, pancreas, and stomach, while relative downregulation is observed in breast, cervix, esophagus, lung, ovary, and uterus.
Survival curves of the clock gene HNF4a in twenty malignant tumors. The survival curves showed that patients with high expression of HNF4a exhibited a significantly longer overall survival (OS, p < 0.05) than patients with low HNF4a expression in BLCA, KIRC, and READ, while patients with high expression of HNF4a exhibited a significantly shorter OS (p < 0.05) than patients with low HNF4a expression in LUSC. No significant difference was observed in other malignant tumors (p > 0.05).
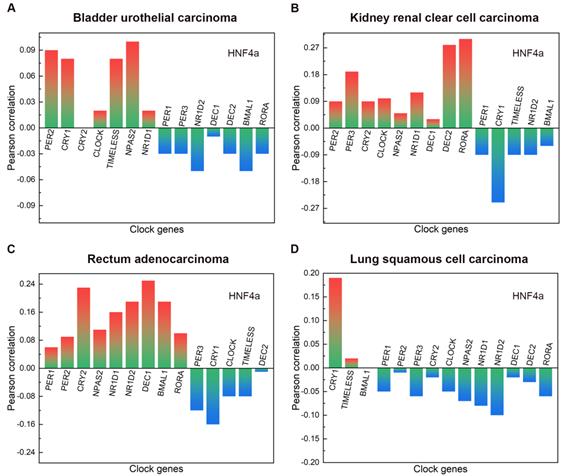
The main clinicopathological features of BLCA are given in Table 1. In BLCA, the high expression rate of HNF4a in the stage group (I+II) was 65.38% (85/130), and that in the stage group (III+IV) was 42.86% (117/273). The high expression rate of HNF4a in the T stage group (T1+T2) was 61.98% (75/121), and that in the T stage group (T3+T4) was 43.43% (109/251). The high expression rate of HNF4a in the lymph node status group (N0) was 53.62% (126/235), and that in the lymph node status group (N1-3) was 39.06% (50/128). Overall, the expression of HNF4a correlated with the stage, T stage, and lymph node status (p < 0.05; Table 2) but not with gender, age, metastasis, and radiation (p > 0.05). As shown in Figure 2A, HNF4a has a correlation with other clock genes in BLCA, which is reflected in the fact that HNF4a correlates positively with PER2 (P = 0.09), CRY1 (P = 0.08), CLOCK (P = 0.02), TIMELESS (P = 0.08), NPAS2 (P = 0.1), and NR1D1 (P = 0.02) and negatively with PER1 (P = -0.03), PER3 (P = -0.03), NR1D2 (P = -0.05), DEC1 (P = -0.01), DEC2 (P = -0.03), BMAL1 (P = -0.05), and RORA (P = -0.03).
The clinical characteristics of bladder urothelial carcinoma.
| Variables | Case, n (%) |
|---|---|
| Age (years) | |
| <60 | 87 (21.48%) |
| ≥60 | 317 (78.27%) |
| NA | 1 (0.25%) |
| Stage | |
| I + II | 130 (32.10%) |
| III + IV | 273 (67.41%) |
| NA | 2 (0.49%) |
| T stage | |
| T1 + T2 | 121 (29.87%) |
| T3 + T4 | 251 (61.98%) |
| TX | 1 (0.25%) |
| NA | 32 (7.90%) |
| Lymph node status | |
| N0 | 235 (58.02%) |
| N1-3 | 128 (31.61%) |
| NX | 36 (8.89%) |
| NA | 6 (1.48%) |
| Metastasis | |
| M0 | 195 (48.15%) |
| M1 | 11 (2.72%) |
| MX | 196 (48.39%) |
| NA | 3 (0.74%) |
| Gender | |
| Female | 106 (14.67%) |
| Male | 299 (85.33%) |
| Radiation | |
| No | 359 (67.39%) |
| Yes | 20 (22.83%) |
| NA | 26 (9.78%) |
NA, not available.
The correlation between HNF4a and other clock genes in BLCA, KIRC, READ, and LUSC. A. In BLCA, HNF4a correlated positively with PER2, CRY1, CLOCK, TIMELESS, NPAS2, and NR1D1 and negatively with PER1, PER3, NR1D2, DEC1-2, BMAL1, and RORA. B. In KIRC, HNF4a correlated positively with PER2-3, CRY2, CLOCK, NPAS2, NR1D1, DEC1-2, and RORA and negatively with PER1, CRY1, TIMELESS, NR1D2, and BMAL1. C. In READ, HNF4a correlated positively with PER1-2, CRY2, NPAS2, NR1D1-2, DEC1, BMAL1, and RORA and negatively with PER3, CRY1, CLOCK, TIMELESS, and DEC2. D. In LUSC, HNF4a correlated positively with CRY1 and TIMELESS and negatively with PER1-3, CRY2, CLOCK, NPAS2, NR1D1-2, DEC1-2, and RORA.
The clinicopathological correlation of HNF4a expression in bladder urothelial carcinoma.
| Variables | HNF4a | χ2 | p-values a | |
|---|---|---|---|---|
| Low expression | High expression | |||
| Gender | 1.761 | 0.185 | ||
| Female | 59 | 47 | ||
| Male | 144 | 155 | ||
| Age (years) | 2.476 | 0.116 | ||
| <60 | 37 | 50 | ||
| ≥60 | 165 | 152 | ||
| Stage | 17.877 | 0.000* | ||
| I + II | 45 | 85 | ||
| III + IV | 156 | 117 | ||
| T stage | 11.247 | 0.001* | ||
| T1 + T2 | 46 | 75 | ||
| T3 + T4 | 142 | 109 | ||
| Lymph node status | 7.028 | 0.008* | ||
| N0 | 109 | 126 | ||
| N1-3 | 78 | 50 | ||
| Metastasis | 1.604 | 0.205 | ||
| M0 | 86 | 109 | ||
| M1 | 7 | 4 | ||
| Radiation | 3.246 | 0.072 | ||
| No | 182 | 177 | ||
| Yes | 6 | 14 | ||
a Pearson Chi-square. * Indicates p < 0.05.
The main clinical and biological features of KIRC are given in Table 3. In KIRC, the high expression rate of HNF4a in the female group was 40.96% (77/188), and that in the male group was 54.78% (189/345). Overall, the expression of HNF4a correlated with gender (p < 0.05; Table 4) but not with age, stage, T stage, lymph node status, metastasis, and radiation (p > 0.05). As shown in Figure 2B, HNF4a has a correlation with other clock genes in KIRC, which is reflected in the fact that HNF4a correlates positively with PER2 (P = 0.09), PER3 (P = 0.19), CRY2 (P = 0.09), CLOCK (P = 0.1), NPAS2 (P = 0.05), NR1D1 (P = 0.12), DEC1 (P = 0.03), DEC2 (P = 0.28), and RORA (P = 0.3) and negatively with PER1 (P = -0.09), CRY1 (P = -0.25), TIMELESS (P = -0.09), NR1D2 (P = -0.09), and BMAL1 (P = -0.06).
The clinical characteristics of kidney renal clear cell carcinoma.
| Variables | Case, n (%) |
|---|---|
| Age (years) | |
| <60 | 245 (45.97%) |
| ≥60 | 287 (53.85%) |
| NA | 1 (0.18%) |
| Stage | |
| I + II | 324 (60.79%) |
| III + IV | 207 (38.84%) |
| NA | 2 (0.37%) |
| T stage | |
| T1 + T2 | 342 (64.17%) |
| T3 + T4 | 191 (35.83%) |
| Lymph node status | |
| N0 | 240 (45.03%) |
| N1-3 | 16 (3.00%) |
| NX | 277 (51.97%) |
| Metastasis | |
| M0 | 422 (79.17%) |
| M1 | 79 (14.82%) |
| MX | 30 (5.63%) |
| NA | 2 (0.38%) |
| Gender | |
| Female | 188 (14.67%) |
| Male | 345 (85.33%) |
| Radiation | |
| No | 141 (26.45%) |
| Yes | 2 (0.38%) |
| NA | 390 (73.17%) |
NA, not available.
The clinicopathological correlation of HNF4a expression in kidney renal clear cell carcinoma.
| Variables | HNF4a | χ2 | p-values a | |
|---|---|---|---|---|
| Low expression | High expression | |||
| Gender | 9.304 | 0.002* | ||
| Female | 111 | 77 | ||
| Male | 156 | 189 | ||
| Age (years) | 0.475 | 0.491 | ||
| <60 | 119 | 126 | ||
| ≥60 | 148 | 139 | ||
| Stage | 2.184 | 0.139 | ||
| I + II | 154 | 170 | ||
| III + IV | 112 | 95 | ||
| T stage | 1.304 | 0.253 | ||
| T1 + T2 | 165 | 177 | ||
| T3 + T4 | 102 | 89 | ||
| Lymph node status | 0.051 | 0.821 | ||
| N0 | 127 | 113 | ||
| N1-3 | 8 | 8 | ||
| Metastasis | 1.534 | 0.215 | ||
| M0 | 203 | 219 | ||
| M1 | 44 | 35 | ||
| Radiation | 1.788 | 0.181 | ||
| No | 74 | 67 | ||
| Yes | 2 | 0 | ||
a Pearson Chi-square. * Indicates p < 0.05.
The main clinical and biological features of READ are given in Table 5. The expression of HNF4a was independent of gender, age, stage, T stage, lymph node status, metastasis, and radiation. (p > 0.05; Table 6). As shown in Figure 2C, HNF4a has a correlation with other clock genes in READ, which is reflected in the fact that HNF4a correlates positively with PER1 (P = 0.06), PER2 (P = 0.09), CRY2 (P = 0.23), NPAS2 (P = 0.11), NR1D1 (P = 0.16), NR1D2 (P = 0.19), DEC1 (P = 0.25), BMAL1 (P = 0.19), and RORA (P = 0.1) and negatively with PER3 (P = -0.12), CRY1 (P = -0.16), CLOCK (P = -0.08), TIMELESS (P = -0.08), and DEC2 (P = -0.01).
The clinical characteristics of rectum adenocarcinoma.
| Variables | Case, n (%) |
|---|---|
| Age (years) | |
| <60 | 41 (42.27%) |
| ≥60 | 56 (57.73%) |
| Stage | |
| I + II | 41 (42.27%) |
| III + IV | 47 (48.45%) |
| NA | 9 (9.28%) |
| T stage | |
| T1 + T2 | 18 (18.56%) |
| T3 + T4 | 77 (79.38%) |
| NA | 2 (2.06%) |
| Lymph node status | |
| N0 | 43 (44.33%) |
| N1-3 | 50 (51.55%) |
| NX | 2 (2.06%) |
| NA | 2 (2.06%) |
| Metastasis | |
| M0 | 67 (69.07%) |
| M1 | 13 (13.41%) |
| MX | 14 (14.43%) |
| NA | 3 (3.09%) |
| Gender | |
| Female | 44 (45.36%) |
| Male | 53 (54.64%) |
| Radiation | |
| No | 73 (75.26%) |
| Yes | 11 (11.34%) |
| NA | 13 (13.40%) |
NA, not available.
The clinicopathological correlation of HNF4a expression in rectum adenocarcinoma.
| Variables | HNF4a | χ2 | p-values a | |
|---|---|---|---|---|
| Low expression | High expression | |||
| Gender | 0.099 | 0.752 | ||
| Female | 23 | 21 | ||
| Male | 26 | 27 | ||
| Age (years) | 0.495 | 0.482 | ||
| <60 | 19 | 22 | ||
| ≥60 | 30 | 26 | ||
| Stage | 1.142 | 0.285 | ||
| I + II | 18 | 23 | ||
| III + IV | 26 | 21 | ||
| T stage | 0.329 | 0.566 | ||
| T1 + T2 | 8 | 10 | ||
| T3 + T4 | 40 | 37 | ||
| Lymph node status | 1.766 | 0.184 | ||
| N0 | 19 | 24 | ||
| N1-3 | 29 | 21 | ||
| Metastasis | 0.268 | 0.605 | ||
| M0 | 36 | 31 | ||
| M1 | 8 | 5 | ||
| Radiation | 0.785 | 0.376 | ||
| No | 37 | 36 | ||
| Yes | 4 | 7 | ||
a Pearson Chi-square. * Indicates p < 0.05.
The main clinical and biological features of LUSC are given in Table 7. In LUSC, the low expression rate of HNF4a in the age group (<60 years) was 62.22% (56/90), and that in the age group (≥60 years) was 46.62% (186/399). Overall, the expression of HNF4a correlated with age (p < 0.05; Table 8) but not with gender, stage, T stage, lymph node status, metastasis, and radiation (p > 0.05). As shown in Figure 2D, HNF4a has a correlation with other clock genes in LUSC, which is reflected in the fact that HNF4a correlates positively with CRY1 (P = 0.19) and TIMELESS (P = 0.02) and negatively with PER1 (P = -0.05), PER2 (P = -0.01), PER3 (P = -0.06), CRY2 (P = -0.02), CLOCK (P = -0.05), NPAS2 (P = -0.07), NR1D1 (P = -0.08), NR1D2 (P = -0.1), DEC1 (P = -0.02), DEC2 (P = -0.03), and RORA (P = -0.06).
The clinical characteristics of lung squamous cell carcinoma.
| Variables | Case, n (%) |
|---|---|
| Age (years) | |
| <60 | 90 (18.18%) |
| ≥60 | 399 (80.61%) |
| NA | 6 (1.21%) |
| Stage | |
| I + II | 90 (18.18%) |
| III + IV | 401 (81.01%) |
| NA | 4 (0.81%) |
| T stage | |
| T1 + T2 | 402 (81.21%) |
| T3 + T4 | 93 (18.79%) |
| Lymph node status | |
| N0 | 316 (58.02%) |
| N1-3 | 173 (31.61%) |
| NX | 6 (8.89%) |
| Metastasis | |
| M0 | 407 (82.22%) |
| M1 | 7 (1.41%) |
| MX | 77 (15.56%) |
| NA | 4 (0.81%) |
| Gender | |
| Female | 129 (26.06%) |
| Male | 366 (73.94%) |
| Radiation | |
| No | 378 (76.36%) |
| Yes | 53 (10.71%) |
| NA | 64 (12.93%) |
NA, not available.
The clinicopathological correlation of HNF4a expression in lung squamous cell carcinoma.
| Variables | HNF4a | χ2 | p-values a | |
|---|---|---|---|---|
| Low expression | High expression | |||
| Gender | 0.007 | 0.935 | ||
| Female | 67 | 67 | ||
| Male | 179 | 182 | ||
| Age (years) | 7.154 | 0.007* | ||
| <60 | 56 | 34 | ||
| ≥60 | 186 | 213 | ||
| Stage | 0.198 | 0.656 | ||
| I + II | 202 | 199 | ||
| III + IV | 43 | 47 | ||
| T stage | 0.168 | 0.682 | ||
| T1 + T2 | 198 | 204 | ||
| T3 + T4 | 48 | 45 | ||
| Lymph node status | 0.093 | 0.760 | ||
| N0 | 158 | 158 | ||
| N1-3 | 84 | 89 | ||
| Metastasis | 0.077 | 0.781 | ||
| M0 | 211 | 196 | ||
| M1 | 4 | 3 | ||
| Radiation | 0.739 | 0.390 | ||
| No | 195 | 183 | ||
| Yes | 24 | 29 | ||
a Pearson Chi-square. * Indicates p < 0.05.
Discussion
The circadian clock exists extensively in various organs and tissues of mammals so that the organism can have a better balance with the rhythm of nature, which has an important biological significance. Mammalian circadian clocks are composed of the master clock and the peripheral clock [29]. Under physiological or pathological stimulations, the body can coordinate the output rhythm of the circadian clock driven by suprachiasmatic nucleus and be transmitted finally to the peripheral clock of the peripheral tissues through a series of signal transduction. The peripheral clock can also regulate the rhythmic change of the local circadian clock of the tissues according to the external stimulation and changes to its own microenvironment.
The master clock of mammals consists primarily of two interlocking transcription/translation circuits. The biological clock molecular regulation system in mammals is a negative feedback transcription/translation circuit formed by more than ten biological clock genes, including BMAL1, CLOCK, CRY, PER, Rev-erbs, Rorα, etc. BMAL1 can bind to the CLOCK protein to form a BMAL1/CLOCK heterodimer, then enter the nucleus and bind to the E-box element of the upstream promoter of the PER and CRY genes and activate its transcription. The proteins of PER and CRY in the cytoplasm can be transferred to the nucleus after continuous accumulation, and combined with the D-box elements in the promoter regions of BMAL1 and CLOCK to exert a negative regulatory function and inhibit the transcriptional activity of BMAL1 and CLOCK [30, 31]. At the same time, CLOCK and BMAL1 heterodimer can also bind to the E-box element on the promoter of the nuclear receptor gene Rev-erbα/β and RORα/β to promote its gene transcription and start the auxiliary circuit of the clock gene transcription/translation oscillation mechanism. Rev-erbα/β and RORα/β proteins competitively bind to receptor response elements in the promoter region of BMAL1. The RORα/β protein promotes the transcription of BMAL1, while the Rev-erbα/β protein inhibits the transcription of BMAL1, thus producing another level of regulation on the main circuit of circadian rhythm [32-34]. The circadian clock regulates the activities of cells, tissues, and organs by regulating many downstream genes to achieve the physiological, biochemical, and complex transformation processes of the body.
HNF4a belongs to the transcription factor of nuclear hormone receptor superfamily 2A, which is mainly found in liver, kidney, and intestinal tissues [35-38]. In lung tissue, the content of HNF4a is low and its positive expression can even be used as a useful marker for difficult diagnosis in small biopsy samples [39]. The analysis results of HNF4a in the GTEx database also showed consistency. HNF4a was highly expressed in bladder, kidney, and rectum but minimally expressed in the lung under normal circumstances. The expression level or the transcriptional activity of various nuclear receptors (NRs) is related closely to the circadian rhythm. Qu et al. showed that HNF4A can effectively inhibit the transcriptional activity of the CLOCK: BMAL1 heterodimer independently of the indirect action of CRY bridging. Moreover, the authors confirmed that when HNF4a is expressed ectopically, the intrinsic circadian clock can change significantly, indicating the importance of HNF4a in the circadian rhythm of its endogenously expressed cells [40]. Caratti's findings revealed that the effect of the glucocorticoid receptor on the energy metabolism and regulation of immunity is felt through the regulation of the HNF4A/HNF6 binding complex by the circadian clock gene NR1D1 in the liver [41]. By analyzing the correlation between clock genes, we found that HNF4A correlated negatively with BMAL1 and positively with NR1D1 in BLCA, which was consistent with previous studies.
Bladder cancer is the ninth most commonly diagnosed cancer in the world. About 80% to 90% of bladder cancers are BLCA [42]. The pathogenesis of bladder cancer is complicated and related to multiple genes [43, 44]. Urine concentration and excretion are regulated by a number of factors, such as vasopressin, thyroxine, and insulin and exhibit distinct circadian rhythms. Circadian clock genes certainly play an important regulatory role in the urinary system, and the malfunctioning of the circadian rhythm could be a contributing mechanism to dysregulation during the development of BLCA. A study by Litlekalsoy et al. analyzed twenty-seven paired tumor and benign tissue samples collected from patients undergoing cystectomy and found that both the oncogene overexpression and reduced transcription of tumor suppressor genes were associated with levels of clock genes [45]. In our study, the HNF4a protein expression in BLCA tissues correlated significantly with the tumor pathology (TNM) grade of the clinicopathological data. The higher the T, N stage, the lower the expression level of HNF4a. The malignant degree of the tumor correlated negatively with the expression level of HNF4a, which suggests that HNF4a plays an important negative role in the development of BLCA. We hypothesize that one aspect may be due to declining transcriptional translation levels of the downstream genes associated with cancer caused directly by HNF4a or that HNF4a reverses the abnormal proliferation and differentiation of cells directly. On the other hand, HNF4a may affect the expression levels of other important circadian clock genes, such as elevated BMAL1 levels, which indirectly affect the regulatory genes and cell signaling pathways involved in DNA damage repair, inflammatory response, cellular metabolism, etc., which can enable cancer cells to avoid deterioration and invasion easily; however, the specific molecular mechanism needs to be researched further. There were only 11 cases (5.34%) with distant metastasis in this study. There was no correlation between the M stage and HNF4a expression level, so we speculated that most of the specimens originated from patients undergoing total cystectomy, but patients with distant metastases of the BLCA generally do not undergo total cystectomy. Differences in survival time between patients with different expressions are also seen in the analysis of the overall survival curve. The overall survival time of patients with high expression of the HNF4a protein is long, whereas, the overall survival time of patients with low HNF4a protein expression is short. Therefore, we hypothesize that HNF4A plays a central role in cell-autonomous circadian rhythm oscillation in a tissue-specific manner in BLCA, pointing to its role in the regulation of the circadian clock network in peripheral tissues.
Non-small cell lung cancer (NSCLC) can be classified as nonsquamous carcinoma, including adenocarcinoma (40% of NSCLC cases) and squamous NSCLC (30% of NSCLC cases) [46]. With the application of vascular-related factor inhibitors, the prognosis of patients with advanced lung adenocarcinoma has improved significantly. However, the performance of these specific inhibitors in patients with squamous NSCLC is poor. Therefore, understanding and studying the potential molecular targets of squamous NSCLC may open new avenues. It has been reported that HNF4a can regulate the development of human mucinous lung adenocarcinoma by inducing MUC3 [47]. Currently, there are no studies on HNF4a and LUSC. Our study showed that the higher the level of HNF4a expression in LUSC, the worse the patient's prognosis, which is significantly different from the results in BLCA, KIRC, and READ. However, the correlation between HNF4a and clinical pathology is not obvious. The specific mechanism of action of HNF4a in LUSC is worthy of our subsequent research and discussion.
The evaluation of the expression of a single clock gene is limited. The studies on transcription data between the different clock genes have shed light on the question of whether the clock-work acts as a system and not only by single gene aberrations. In different tumors, the mutual influence of different clock genes leads to different outcomes, which reminds us that when we conduct basic research on a single clock gene, we should pay attention to the promotion or inhibition influence of other clock genes. It is the regulation of these systems that makes the circadian rhythm so orderly, and even forms characteristic asynchronous circadian rhythms in cancers.
In conclusion, this study shows that HNF4a could be a tumor suppressor gene, and its loss of expression in BLCA tissues is likely to promote the occurrence and progression of BLCA. The high expression of HNF4a in the bladder mucosa of patients may help to inhibit the occurrence and development of tumors and could have a certain guiding role in the early diagnosis and treatment of patients. The HNF4a protein could be used as one of the prognostic indicators for patients with BLCA, and it could become a possible alternative target for BLCA treatment. The combined detection of the HNF4a protein and other factors could improve the prediction of the overall survival of patients with BLCA greatly and provide a basis for early detection and early clinical treatment intervention. Therefore, we intend to explore the related mechanism of the HNF4a gene's low-expression in BLCA further and hope to provide a clinical basis for the early diagnosis and prognosis of bladder cancer patients.
Supplementary Material
Supplementary figure S1.
Acknowledgements
Funding
This study was supported by the National Key Research and Development Program of China (2018YFC2002000), the National Natural Science Foundation of China (81702885), the National Natural Science Foundation of China (81874232).
Authors' contributions
Zhi-fan Xiong and Sheng-li Yang conceived and designed the experiments. Meng-jun Qiu, Li Zhang, and Xie-fan Fang carried out the research. Qiu-ting Li, Li-sheng Zhu, and Bin Zhang contributed the analysis tools and analyzed the data for the study. Meng-jun Qiu wrote the paper. All authors have read and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Watt AJ FAU Garrison WD, Garrison WD FAU Duncan SA, Duncan SA. HNF4: a central regulator of hepatocyte differentiation and function. Hepatology. 2003;37(6):1249-53
2. Tirona RG, Lee W FAU Leake BF, Leake BF FAU Lan L, Lan LB FAU Cline CB, Cline CB FAU Lamba V, Lamba V FAU Parviz F. et al. The orphan nuclear receptor HNF4alpha determines PXR- and CAR-mediated xenobiotic induction of CYP3A4. Nat Med. 2003;9(2):220-4
3. Inoue Y, Hayhurst GP FAU Inoue J, Inoue J FAU Mori M, Mori M FAU Gonzalez FJ, Gonzalez FJ. Defective ureagenesis in mice carrying a liver-specific disruption of hepatocyte nuclear factor 4alpha (HNF4alpha). HNF4alpha regulates ornithine transcarbamylase in vivo. J Biol Chem. 2002;277(28):25257-65
4. Stoffel M, Duncan SA. The maturity-onset diabetes of the young (MODY1) transcription factor HNF4alpha regulates expression of genes required for glucose transport and metabolism. Proc Natl Acad Sci U S A. 1997;94(24):13209-14
5. Xie X, Liao H FAU Dang H, Dang H FAU Pang W, Pang W FAU Guan Y, Guan Y FAU Wang X, Wang X FAU Shyy JY. et al. Down-regulation of hepatic HNF4alpha gene expression during hyperinsulinemia via SREBPs. Mol Endocrinol. 2009;23(4):434-43
6. Zhang Y, Yin L FAU Anderson J, Anderson J FAU Ma H, Ma H FAU Gonzalez FJ, Gonzalez FJ FAU Willson TM, Willson TM FAU Edwards PA. et al. Identification of novel pathways that control farnesoid X receptor-mediated hypocholesterolemia. J Biol Chem. 2010;285(5):3035-43
7. Chiang JY. Regulation of bile acid synthesis: pathways, nuclear receptors, and mechanisms. J Hepatol. 2004;40(3):539-51
8. Hwang-Verslues WW, Sladek FM. HNF4alpha-role in drug metabolism and potential drug target? Curr Opin Pharmacol. 2010;10(6):698-705
9. Lazarevich NL, Cheremnova OA FAU Varga EV, Varga EV FAU Ovchinnikov DA, Ovchinnikov DA FAU Kudrjavtseva EI, Kudrjavtseva EI FAU Morozova OV, Morozova OV FAU Fleishman DI. et al. Progression of HCC in mice is associated with a downregulation in the expression of hepatocyte nuclear factors. Hepatology. 2004;39(4):1038-47
10. Lucas B, Grigo K FAU Erdmann S, Erdmann S FAU Lausen J, Lausen J FAU Klein-Hitpass L, Klein-Hitpass L FAU Ryffel GU, Ryffel GU. HNF4alpha reduces proliferation of kidney cells and affects genes deregulated in renal cell carcinoma. Oncogene. 2005;24(42):6418-31
11. Spath GF, Weiss MC. Hepatocyte nuclear factor 4 provokes expression of epithelial marker genes, acting as a morphogen in dedifferentiated hepatoma cells. J Cell Biol. 1998;140(4):935-46
12. Chiba H, Itoh T FAU Satohisa S, Satohisa S FAU Sakai N, Sakai N FAU Noguchi H, Noguchi H FAU Osanai M, Osanai M FAU Kojima T. et al. Activation of p21CIP1/WAF1 gene expression and inhibition of cell proliferation by overexpression of hepatocyte nuclear factor-4alpha. Exp Cell Res. 2005;302(1):11-21
13. Logan RW, McClung CA. Rhythms of life: circadian disruption and brain disorders across the lifespan. Nat Rev Neurosci. 2019;20(1):49-65
14. Panda S. Circadian physiology of metabolism. Science. 2016;354(6315):1008-1015
15. Sato TK, Panda S FAU Miraglia LJ, Miraglia LJ FAU Reyes TM, Reyes TM FAU Rudic RD, Rudic RD FAU McNamara P, McNamara P FAU Naik KA. et al. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron. 2004;43(4):527-37
16. Chen L, Yang G. Recent advances in circadian rhythms in cardiovascular system. Front Pharmacol. 2015;6:71
17. Hastings MH, Goedert M. Circadian clocks and neurodegenerative diseases: time to aggregate? Curr Opin Neurobiol. 2013;23(5):880-7
18. Marcheva B, Ramsey KM FAU Buhr ED, Buhr ED FAU Kobayashi Y, Kobayashi Y FAU Su H, Su H FAU Ko CH, Ko CH FAU Ivanova G. et al. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature. 2010;466(7306):627-31
19. Qiu MJ, Liu LP, Jin S, Fang XF, He XX, Xiong ZF. et al. Research on circadian clock genes in common abdominal malignant tumors. Chronobiol Int. 2019;36(7):906-918
20. Altman BJ, Hsieh AL, Sengupta A, Krishnanaiah SY, Stine ZE, Walton ZE. et al. MYC Disrupts the Circadian Clock and Metabolism in Cancer Cells. Cell Metab. 2015;22(6):1009-19
21. Bonzo JA, Ferry CH FAU Matsubara T, Matsubara T FAU Kim J, Kim JH FAU Gonzalez FJ, Gonzalez FJ. Suppression of hepatocyte proliferation by hepatocyte nuclear factor 4alpha in adult mice. J Biol Chem. 2012;287(10):7345-56
22. Ning BF, Ding J FAU Yin C, Yin C FAU Zhong W, Zhong W FAU Wu K, Wu K FAU Zeng X, Zeng X FAU Yang W. et al. Hepatocyte nuclear factor 4 alpha suppresses the development of hepatocellular carcinoma. Cancer Res. 2010;70(19):7640-51
23. Hatziapostolou M, Polytarchou C FAU Aggelidou E, Aggelidou E FAU Drakaki A, Drakaki A FAU Poultsides GA, Poultsides GA FAU Jaeger SA, Jaeger SA FAU Ogata H. et al. An HNF4alpha-miRNA inflammatory feedback circuit regulates hepatocellular oncogenesis. Cell. 2011;147(6):1233-47
24. Martinez-Jimenez CP, Kyrmizi I FAU Cardot P, Cardot P FAU Gonzalez FJ, Gonzalez FJ FAU Talianidis I, Talianidis I. Hepatocyte nuclear factor 4alpha coordinates a transcription factor network regulating hepatic fatty acid metabolism. Mol Cell Biol. 2010;30(3):565-77
25. Lee YK, Dell H FAU Dowhan DH, Dowhan DH FAU Hadzopoulou-Cladaras M, Hadzopoulou-Cladaras M FAU Moore DD, Moore DD. The orphan nuclear receptor SHP inhibits hepatocyte nuclear factor 4 and retinoid X receptor transactivation: two mechanisms for repression. Mol Cell Biol. 2000;20(1):187-95
26. Ruse MJ, Privalsky ML FAU Sladek FM, Sladek FM. Competitive cofactor recruitment by orphan receptor hepatocyte nuclear factor 4alpha1: modulation by the F domain. Mol Cell Biol. 2002;22(6):1626-38
27. Fekry B, Ribas-Latre A, Baumgartner C, Deans JR, Kwok C, Patel P. et al. Incompatibility of the circadian protein BMAL1 and HNF4alpha in hepatocellular carcinoma. Nat Commun. 2018;9(1):4349
28. Yao HS, Wang J, Zhang XP, Wang LZ, Wang Y, Li XX. et al. Hepatocyte nuclear factor 4alpha suppresses the aggravation of colon carcinoma. Mol Carcinog. 2016;55(5):458-72
29. Mohawk JA, Takahashi JS. Cell autonomy and synchrony of suprachiasmatic nucleus circadian oscillators. Trends Neurosci. 2011;34(7):349-58
30. Huang N, Chelliah Y FAU Shan Y, Shan Y FAU Taylor CA, Taylor CA FAU Yoo S, Yoo SH FAU Partch C, Partch C FAU Green CB. et al. Crystal structure of the heterodimeric CLOCK: BMAL1 transcriptional activator complex. Science. 2012;337(6091):189-94
31. Casper RF, Gladanac B. Introduction: circadian rhythm and its disruption: impact on reproductive function. Fertil Steril. 2014;102(2):319-20
32. Ikeda R, Tsuchiya Y, Koike N. et al. REV-ERBα and REV-ERBβ function as key factors regulating Mammalian Circadian Output. Sci Rep. 2019;9(1):10171
33. Liu AC, Tran HG, Zhang EE, Priest AA, Welsh DK, Kay SA. Redundant function of REV-ERBalpha and beta and non-essential role for Bmal1 cycling in transcriptional regulation of intracellular circadian rhythms. PLoS Genet. 2008;4(2):e1000023
34. Guillaumond F, Dardente H, Giguère V, Cermakian N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J Biol Rhythms. 2005;20(5):391-403
35. Saandi T, Baraille F, Derbal-Wolfrom L. et al. Regulation of the tumor suppressor homeogene Cdx2 by HNF4α in intestinal cancer. Oncogene. 2013;32(32):3782-8
36. Ji D, Chen GF, Wang JC. et al. Identification of TAF1, HNF4A, and CALM2 as potential therapeutic target genes for liver fibrosis. J Cell Physiol. 2019;234(6):9045-9051
37. Chen L, Vasoya RP, Toke NH. et al. HNF4 Regulates Fatty Acid Oxidation and Is Required for Renewal of Intestinal Stem Cells in Mice. Gastroenterology. 2020;158(4):985-999.e9
38. Martovetsky G, Bush KT, Nigam SK. Kidney versus Liver Specification of SLC and ABC Drug Transporters, Tight Junction Molecules, and Biomarkers. Drug Metab Dispos. 2016;44(7):1050-60
39. Sugano M, Nagasaka T, Sasaki E. et al. HNF4α as a marker for invasive mucinous adenocarcinoma of the lung. Am J Surg Pathol. 2013;37(2):211-8
40. Qu M, Duffy T, Hirota T, Kay SAO. Nuclear receptor HNF4A transrepresses CLOCK: BMAL1 and modulates tissue-specific circadian networks. Proc Natl Acad Sci U S A. 2018;115(52):E12305-E12312
41. Caratti G, Iqbal M, Hunter L, Kim D, Wang P, Vonslow RM. et al. REVERBa couples the circadian clock to hepatic glucocorticoid action. J Clin Invest. 2018;128(10):4454-4471
42. Antoni S, Ferlay J, Soerjomataram I, Znaor A, Jemal A, Bray F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur Urol. 2017;71(1):96-108
43. Jacobs BL, Lee CT FAU Montie JE, Montie JE. Bladder cancer in 2010: how far have we come? CA Cancer J Clin. 2010;60(4):244-72
44. Fujimoto K, Yamada Y FAU Okajima E, Okajima E FAU Kakizoe T, Kakizoe T FAU Sasaki H, Sasaki H FAU Sugimura T, Sugimura T FAU Terada M. et al. Frequent association of p53 gene mutation in invasive bladder cancer. Cancer Res. 1992;52(6):1393-8
45. Litlekalsoy JAO, Rostad K, Kalland KH, Hostmark JG, Laerum OD. Expression of circadian clock genes and proteins in urothelial cancer is related to cancer-associated genes. BMC Cancer. 2016;16:549
46. Piperdi B, Merla A FAU Perez-Soler R, Perez-Soler R. Targeting angiogenesis in squamous non-small cell lung cancer. Drugs. 2014;74(4):403-13
47. Guo M, Tomoshige K, Meister M, Muley T, Fukazawa T, Tsuchiya T. et al. Gene signature driving invasive mucinous adenocarcinoma of the lung. EMBO Mol Med. 2017;9(4):462-481
Author contact
![]() Corresponding authors: Dr. Sheng-li Yang, Cancer Center, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China, E-mail: yangshengliedu.cn. And to Prof. Zhi-fan Xiong, Division of Gastroenterology, Liyuan Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430077, China. E-mail: xiongzhifancom.
Corresponding authors: Dr. Sheng-li Yang, Cancer Center, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China, E-mail: yangshengliedu.cn. And to Prof. Zhi-fan Xiong, Division of Gastroenterology, Liyuan Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430077, China. E-mail: xiongzhifancom.